Immunbiologie
Die Immunbiologie liefert Erklärungen über:
den Aufbau der Immunreaktion,
den Aufbau des Immunsystems und seine genetischen Grundlagen,
Anwendungen bei der Schutzimpfung und Organtransplantation,
die Immunschwäche - Krankheit AIDS.
Das Immunsystem des Menschen
In den menschlichen Körper gelangen ständig zahlreiche Krankheitserreger, vor allem Bakterien und Viren. Dennoch kommt es bei den meisten Menschen nur selten zu einer Erkrankung. Das erste Hindernis gegen das Eindringen von Fremdkörpern bilden die Haut und die Schleimhäute. Zweitens verfügt der Körper über ein komplexes Abwehrsystem. Dieses ist gegen viele Krankheitserreger gleichermaßen wirksam und wird unter dem Begriff unspezifische Abwehr oder Resistenz zusammengefaßt. Drittens verfügt der Körper über ein Immunsystem: Dringen Mikroorganismen, vor allem über kleine Verletzungen, in den Bereich von Gewebeflüssigkeit und Kreislaufsystem vor, so erkennen bestimmte Zellen die eingedrungenen Fremdkörper und lösen eine spezifische Reaktion aus, welche die Erreger unschädlich macht. Diese Reaktion nennt man Immunreaktion. Der Körper kann dabei also zwischen "fremd" und "eigen" unterscheiden.
Unspezifische Abwehr und Immunität
Unspezifische Abwehr: Resistenz
Zellen der Schleimhäute bilden schleimige Sekrete, die Mikroorganismen und andere Fremdkörper einhüllen, sodaß sie zusammen mit dem Schleim aus dem Körper entfernt werden können. Tränenflüssigkeit und Nasensekret enthalten das von dem sekretbildenden Zellen abgegebene Enzym Lysozym, welches die Zellwände eindringender Bakterien zerstört. Im Körperinneren gibt es ebenfalls Stoffe, die gegen Mikroorganismen und Viren wirken. Interferone werden von Zellen gebildet, die von Viren befallen worden sind. Dies sind Proteine, welche in den Zellen die Bildung schützender, gegen Viren gerichteter Proteine auslösen und außerdem auf Zellen des Immunsystems einwirken. Im Blutserum befinden sich ferner Proteine (Enzyme und Enzymvorstufen), die in komplizierter Weise als Komplementsystem zusammenarbeiten. Sie zerstören die Membranen von Mikroorganismen, spalten Proteine und stimulieren die Phagozytose(siehe unten) durch Makrophagen. Die Bezeichnung leitet sich davon ab, daß diese System die Immunreaktion ergänzt (complement = Ergänzung).
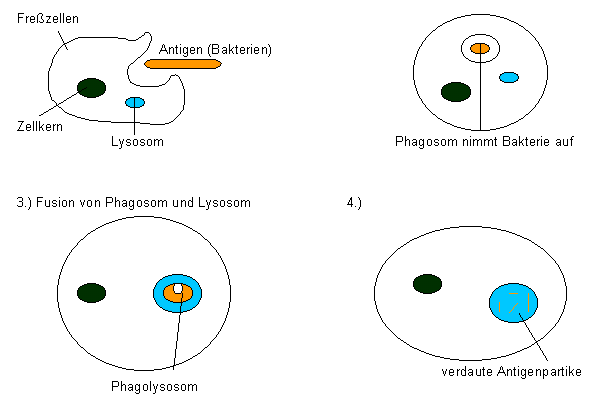
Zur unspezifischen Resistenz tragen auch bestimmte Weiße Blutzellen (Leukozyten) bei, die eingedrungene Fremdkörper durch Phagozytose aufnehmen und intrazellulär abbauen. Unter den Leukozyten sind solche mit großen, gelappten Zellkernen und körnchenförmigen (granulären) Einschlüssen im Cytoplasma (Granulozyten); sie zirkulieren im Blut und gelangen auf diese Weise rasch an die Orte, an denen sie benötigt werden. Andere, Makrophagen genannt, finden sich vor allem in den Lymphknoten, Leber, Milz, Lunge und Nieren. Sie stehen mit den Immunreaktionen in enger Verbindung. Makrophagen nehmen Bakterien durch Phagozytose auf. In den Zellen entstehen dann Vakuolen mit den Bakterien, die dort durch Enzyme der Lysosomen verdaut werden (Unterschied zu Granulozyten). Die Phagozytose wird vom Komplementsystem reguliert. Schon dadurch wird erkennbar, wie vielfältig die einzelnen Teile des Abwehrsystems untereinander in Wechselwirkung stehen.
Letztlich unterliegen alle eingedrungenen Bakterien und Viren der Phagozytose. Hierbei können phagozytierende Zellen zerstört und dann im Eiter ausgeschieden werden. Der Eiter enthält außerdem andere zerstörte Zellen.
1.) Umfließen des Antigens 2.)
Spezifische Abwehr: Immunität
Im Gegensatz zur Resistenz ist die Immunität durch eine hohe Spezifität ausgezeichnet. Das zeigt folgender Versuch: Man injiziert einem Kaninchen eine kleine Menge Bakterien einer bestimmten Art und wiederholt dies im Laufe weniger Wochen mehrmals. Anschließend entnimmt man ihm einige cm3 Blut und gewinnt das Serum daraus. Verdünnt man nun das Serum (im Verhältnis 1: 1000) und mischt es mit einer Kultur der injizierten Bakterienart, so verklumpen die Bakterien. Beim Vermischen der Bakterien mit Serum nicht behandelter Kaninchen tritt kein Verklumpen auf, ebenso wenig beim Vermischen von Bakterien einer anderen Art mit dem Serum des behandelten. Man kann daraus schließen, daß im Körper des Kaninchens Stoffe gebildet werden, die mit der Bakterienzellmembran so reagiert, daß die Bakterien verklumpen. Diese von Körperzellen gebildeten Stoffe heißen Antikörper. Sie werden offenbar gebildet, wenn körperfremde Stoffe- im vorliegenden Fall Teile der Zellmembran des Bakteriums- in die Blutbahn des Kaninchens gelangt sind. Stoffe, die eine Bildung von Antikörpern auslösen, nennt man Antigene. Der Begriff "Antigene" weckt häufig irrtümliche Gedankenverbindungen zu "Gen". Er ist aber verkürzt gebildet aus Anti-somato-gen (= gegen den Körper bildend). Als Antigene können Peptide, Proteine, Polysaccharide und Polynucleotide wirken. Der Antikörper wird dabei nicht gegen das Antigenmolekül als Ganzes gebildet, sondern nur gegen bestimmte Atomanordnungen an der Oberfläche dieser Moleküle. Diese Bereiche nennt man Epitope. Die Antikörper reagieren mit dem Eptiop des Antigens, das die Bildung des betreffenden Antikörpers verursacht hat (Antigen- Antikörper- Reaktion).
Während Residenz auch bei vielen Wirbellosen auftritt, ist die Antikörperbildung bisher nur bei Wirbeltieren festgestellt worden.
Antigene werden von Lymphozyten erkannt. Diese machen etwa 25% der Weißen Blutzellen (Leukozyten) aus. Sie entstehen im Knochenmark, das deshalb als primäres Immunorgan bezeichnet wird. Ist das Antigen erkannt, so reagieren die verschiedenen Arten von Lymphozyten unterschiedlich:
Ungefähr 10% der Lymphozyten erzeugen Antikörper und geben diese an Lymphe und Blut (humorale Bestandteile des Körpers) ab. Diese Antikörper bilden den Zellen reifen im Knochenmark (bone-marrow) und heißen B-Lymphozyten. Nach der Entwicklung wandern die in die sekundären Immunorgane (Lymphknoten, Milz, Wurmfortsatz, Mandeln) ein oder kreisen im Blutstrom. Die sind für die humorale Immunantwort verantwortlich.
Ungefähr 90% der Lymphozyten haben die Aufgabe, Zellen zu zerstören, die Antigene tragen. Ihre Vorstufen wandern aus dem Knochenmark zunächst in den Thymus ein und reifen dort; sie heißen daher T- Lymphozyten. Sie dienen der zellulären Immunantwort.
Der Thymus reicht bei Jugendlichen in Form zweier Lappen im oberen Brustabschnitt von Kehlkopf bis zum Herzen. Beim Erwachsenen ist er rückgebildet. Später gelangen die T- Lymphozyten ebenfalls in die sekundären Immunorgane, wo sie sich festsetzten und weitervermehren. Von dort werden sie auch ins Blut abgegeben. Die meisten T- und B-Lymphozyten, die sich außerhalb der sekundären Immunorgane befinden, wandern zwischen Blut und Lymphe hin und her.
Bestandteile des Immunsystems
Antikörper:
Die Antikörper sind globuläre Proteine. Man sie Immunglobuline (Ig). Sie werden in fünf Klassen eingeteilt; ihre Baueinheiten sind stets gleichartig. Der häufigste und am besten bekannte Typus sind die Immunglobuline G (IgG). IgG-Moleküle bilden 75 bis 80 % der freien Immunglobuline im Organismus. 80 g zellfreies Blutplasma enthält etwa 1 g IgG. Jedes IgG-Molekül hat zwei Bindungsstellen, die so gebaut sind, daß dort nur solche Antigene gebunden werden können, welche die Bindung des betreffenden Antikörpers veranlaßt haben. Ein Anitkörpermolekül verbindet sich also mit zwei Antigen- Molekülen. Besitzt ein Antigen mehr als eine Bindungsstelle für Antikörpermoleküle, so einstehen größere Komplexe (Immunkomplexe). Deren Größe hängt von der relativen Konzentration von Antigen und Antikörper ab. Sie können so groß werden, daß sie nicht mehr löslich sind und dann ausfallen (Präzipitation).
Lymphozyten:
Die Lymphozyten entstehen aus Zellen, die beim Fetus aus der Leber ins Knochenmark wandern und dort zu Stammzellen aller Blutzellen werden.
Wie schon erwähnt erfolgt die Ausdifferenzierung teils im Thymus (T-Zellen), teils im Knochenmark (B-Zellen). Ein erwachsener Mensch besitzt etwa 2.1012 Lymphozyten, die zusammen etwa 1 kg wiegen, sowie etwa 1020 Antikörpermoleküle, die ebenfalls 1 kg des Körpergewichts ausmachen.
B-Lymphozyten
besitzen in ihrer Zellmembran etwa 100 000 Rezeptormoleküle für ein bestimmtes Antigen. Diese Rezeptoren sind gebaut wie Immunglobuline der IgM-Klasse, sind aber Bestandteile der Membran. Jede B-Zelle besitzt nur Rezeptoren mit einer Spezifität, kann also nur ein bestimmtes Antigen erkennen und binden.
Nach der Bindung der Antigen- Moleküle werden diese in die Zelle aufgenommen und intrazellulär gespalten. Teile dieser Moleküle, die noch als Antigenen wirksam sind, werden dann in die Zellmembran der B-Zelle eingesetzt. Die gebundenen Antigene aktivieren T-Helfer-Lymphozyten, die dann ihrerseits B-Lymphozyten zur Teilung anregen.
Nach Aktivierung teilen sich die B-Lymphozyten mehrfach, so daß zahlreiche Zellen gleicher Spezifität entstehen. Diese Tochterzellen differenzieren sich vorwiegend zu Plasmazellen und bilden Antikörper. Von jeder Plasmazelle werden je Sekunde etwa 2000 gleiche Antikörpermoleküle an die Körperflüssigkeit abgegeben. Die Plasmazellen leben nur wenige Wochen. Durch energiereiche Strahlungen werden diese kurzlebigen Zellen geschädigt (Strahlenwirkung auf das Immunsystem). Einige der B-Zellen differenzieren sich zu Gedächtniszellen, die über Jahre hinweg erhalten bleiben. Wenn sie später erneut auf das gleiche Antigen treffen werden sie aktiviert und teilen sich rasch, so daß in viel kürzerer Zeit mehr Antikörper gebildet werden als beim ersten Kontakt. Die zweite Immunreaktion verläuft daher viel rascher und oft heftiger. Gegen verschiedene Erreger bleibt der Mensch deshalb lebenslang immun (z.B.: gegen die Erreger vieler Kinderkrankheiten).
Jeder B-Zell-Klon erkennt nur ein bestimmtes Antigen. Man weiß aber, daß beispielsweise im Körper des Menschen insgesamt gegen Millionen von Antigenen Antikörper gebildet werden können. Also muß es Millionen verschieder B-Zell-Klone im Körper geben.
T-Lymphozyten
erzeugen keine Antikörpermoleküle; sie besitzen aber Membranrezeptoren mit Spezifität für ein Antigen; diese heißen T-Zell-Rezeptoren. Sie erkennen nur Teile von Antigenen, die an die Membranen bestimmter anderer Zellen gebunden sind. Trifft ein T-Lymphozyt ein membrangebundenes Antigen, so wird er aktiviert und vermehrt sich durch Teilung. Dabei werden ebenfalls Gedächtniszellen gebildet. Die T-Lymphozyten haben unterschiedliche Aufgaben, so daß man verschiedenen Arten unterscheiden kann: die wichtigsten sind:
T-Helfer-Zellen; sie stimulieren nach Erkennung eines Antigens die Teilung der entsprechenden B-Zellen zu Plasmazellen und damit die Bildung von Antikörpern.
T-Unterdrücker-Zellen; sie hemmen die Teilung der B-Zellen und die Bildung von T-Killer-Zellen.
T-Killer-Zellen; sie erkennen und vernichten körpereigene Zellen, die von Viren befallen worden sind und körperfremde Zellen. Sie heißen auch cytotoxische T-Zellen.
T-Zellen geben Signalstoffe (Gewebshormone), die auf andere Zellen des Immunsystems wirken. Diese Stoffe heißen Interleukine.
Antigen-Darbietung (Präsentation).
Dringt ein als Antigen wirksamer Fremdkörper in den Organismus ein, so ist eine zu diesem Antigen passende B-Zelle normalerweiser nicht in der Nähe. Dies würde die Immunreaktion verzögern. Daher wird das Antigen zunächst unspezifisch von einem Makrophagen aufgenommen und in dessen Membran gebunden. Der Makrophage tritt dann in einer Art Schlüssel-Schloss-Reaktion mit B- und T-Zellen in Kontakt und 'präsentiert' das Antigen dieser Zellen, d.h. er bringt es mit Rezeptoren auf deren Zellmembran in Verbindung. Man nennt diese Makrophagen daher Antigen-präsentierenden Zellen. In den Lymphknoten sind die Antigen-präsentierenden Zellen so angeordnet, daß die B- und T-Zellen mit ihnen in engen Kontakt gelangen müssen. Zusätzlich erzeugen sie Interleukine, welche T-Lymphozyten zur Teilung anregen. Gleichzeitig entstehen T-Gedächtniszellen. Die T-Helfer-Zellen beschleunigen dann die Teilung von B-Zellen und ihrer Reifung zu Plasmazellen und simulieren ebenfalls T-Killer-Zellen. Mit Verzögerung folgt eine Stimulation von T-Unterdrücker-Zellen, welche ihrerseits die weitere Teilung und Reifung von B-Zellen zu Plasmazellen hemmen.
T-Zell-Rezeptor und MHC-Proteine.
Die T-Lymphozyten erkennen nur membrangebundene Antigene. Dabei reagieren die Rezeptoren der jeweiligen T-Zelle mit einem dargebotenen Teilstück des Antigens. Diese Reaktion kann nur stattfinden, wenn das Antigen-Teilstück auf der darbietenden Zelle an ein Membranprotein gebunden an der äußeren Oberfläche vorliegt. Die T-Zelle erkennt dabei gleichzeitig dieses Mebranprotein. An ihnen erkennt das Immunsystem auch körperfremde Zellen, so daß diese angegriffen werden. Sie werden daher als Gewebsverträtlichkeits-Proteine (kurz MHC-Proteine von major histocompatibility complex) bezeichnet. Beim Menschen wird ihre Bildung von sechs Genen gesteuert. Den zahlreichen möglichen Kombinationen dieser Gene entsprechend gibt es eine große Zahl von Varianten der MHC-Proteine. Daher besitzen zwei nicht sehr nahe verwandte Meschen kaum je den gleichen Satz von MHC-Proteinen. MHC-Proteine, die anders gebaut sind als die körpereigenen, werden von T-Zellen nicht als solche erkannt. Sie sind daher Fremdproteine und wirken als Antigene. An MHC-Proteinen kann das Immunsystem also körpereigene von körperfremden Zellen unterscheiden. Dies ist bei Gewebs und Orgenverpflanzung von Bedeutung; die fremden MHC-Proteine wirken als 'Gewebs-Antigene' und lösen die Bildung von Antikörpern aus, so daß verpflanzte Gewebe abgestoßen werden.
Weitere Zellen des Immunsystems:
An den Immunreaktionen sind weiter Zellen beteiligt. Mastzellen kommen u.a. in der Nähe von Schleimhäuten vor und tragen an ihrer Oberfläche Antikörper der IgE-Gruppe. Sie enthalten in Golgi-Vesikeln Histamin und Serotonin sowie Prostaglandine und Interleukine. Wird das passende Antigen gebunden, so wird der Inhalt der Golgi-Vesikeln freigesetzt. Er verursacht eine Erweiterung der Blutgefäße und verstärkt die Durchblutung des Gewebes, das sich dadurch rötet. Außerdem häufen sich Lymphozyten und Freßzellen an, so daß eine intensive Immunreaktion abläuft, die als lokale Entzündungsreaktion erkennbar wird.
Bestimmt Granulozyten töten solche Fremdorganismen ab, die für die Aufnahme durch Makrophagen zu groß sind. Ferner gibt es Zellen, die Fremdorganismen ohne die Mitwirkung von Antikörper erkennen und vernichten. Sie greifen sogar körpereigenen Zellen an, wenn deren Oberfläche verändert ist. Diese natürlichen Killerzellen sind wahrscheinlich verantwortlich für die rasche Vernichtung neu entstandener Krebszellen, da bei diesen die Zellmembran Veränderungen aufweist.
Immunreaktion
Antigen-Antikörper-Reaktion
Kommt ein Antigen erstmals in den Körper, so läuft die Immunreaktion in 3 Phasen ab. In der ersten, der Erkennungsphase, muß das Antigen zunächst gebunden und einigen T-Lymphozyten präsentiert werden. Diese werden dadurch aktiviert und zur Teilung angeregt. Es entstehen vermehrt T-Helfer-Zellen; sie lösen bei denjenigen B-Lymphozyten Teilung aus, die Antikörper gegen das gleiche Antigen bilden können.
In der zweiten Phase, der Differenzierungsphase, vermehren sich die B-Lymphozyten stark und differenzieren sich dabei hauptsächlich zu Plasmazellen.; einige werden zu Gedächtniszellen. Die starke Vermehrung der Lymphozyten ist als Schwellung der Lymphknoten in der Nähe eines Infektionsherdes spürbar.
In der dritten Phase, der Wirkungsphase, erfolgt die Antigen-Antikörper-Reaktion und es entsteht der Immunkomplex
Immunkomplexe können das Komplementsystem aktivieren, dessen Proteine dann eine ganze Abfolge von Reaktionen auslösen. Dazu gehört der Abbau von Fremdproteinen, eine Stimulierung der phagozytierenden Zellen und eine chemotaktische Anlockung weiterer solcher Zellen. Diese nehmen dann die Immunkomplexe auf und bauen sie ab. Ist der Abbau verzögert, so kommt es zu Störungen in Form von Allergien.
Immunoglobuline können sich an die Oberfläche eines aus einer Zelle ragenden Antigens (z.B. einer Bakterienzellwand) anheften und die Zellen, die viele Antigene besitzen, verkleben (Agglutination).
Zugleich erhöht sich die Durchlässigkeit der Kapillarwände für Proteine, sodaß vermehrt Antikörper aus dem Blut ins Gewebe gelangen. Infolgedessen tritt auch mehr Flüssigkeit aus den Kapillaren aus, die Infektionsstelle schwillt an
In der vierten Phase, der sogenannten Abschaltphase, hört infolge der Wirkung der T-Unterdrücker-Zellen die Immunreaktion allmählich auf. Wenn kein Antigen mehr vorhanden ist, werden auch keine neuen Antikörper mehr gebildet.
Infektionen
Infektionen werden durch Mikroorganismen, vor allem Bakterien, und durch Viren verursacht. Eine Infektion des Organismus muß allerdings nicht immer zu einer Erkrankung führen; so sind manche Personen zwar Träger und Verbreiter von Krankheitserregern, selbst aber nicht krank.
Immunreaktion gegen Bakterien
Sie hängt davon ab, ob sich die Bakterien außerhalb der Zellen oder intrazellulär vermehren. Bei extrazellulärer Vermehrung führen die Antigene der Bakterienoberfläche rasch zu den geschilderten Immunreaktionen. Geben die Bakterien giftige Stoffe (Toxine) ab, so werden diese als freie Antigene von Antikörpern gebunden und unschädlich gemacht, sofern die Giftproduktion nicht zu rasch zunimmt. Die an die Bakterienoberfläche gebundene Antikörper verursachen zusätzlich einen Angriff des Komplementsystems, welches die Bakterienmembran auflösen kann.
Schleimhäute geben im Sekret größere Menge von Antikörpern der IgA-Gruppe ab, wenn B-Lymphozyten zu deren Bildung stimuliert sind. Die Antikörper binden Bakterien, sodaß diese die Schleimhautzellen nicht mehr angreifen können. IgA-Moleküle treten auch in der Muttermilch auf; die Antikörper bildenden Zellen liegen in den Lymphknoten der Brustdrüse. Sie stammen aus Lymphknoten des Darmbereichs, wo sie Antikörper "kennen gelernt" haben. Die IgA-Antikörper schützen daher den Säugling bei Darminfektionen.
Vermehren sich Bakterien intrazellulär (z.B. Tuberkulosebakterien), so werden die befallenen Zellen von T-Killer-Zellen angegriffen, weil die Membran solcher Zellen veränderte Proteine aufweist. Diese Form der Immunreaktion erfolgt allerdings langsamer.
Gegen Tuberkulosebakterien werden die meisten Menschen immun. Daß sie es sind, zeigt der Tuberkulin-Hauttest, bei dem Tuberkulin, ein Extrakt aus Tuberkelbakterien, als Antigen in die Haut eingerieben wird. In der Regel kommt es dort im Verlauf von 2-4 Tagen zu einer schwachen Entzündungsreaktion (Rötung). Die dort entstehenden Immunkomplexe aktivieren nämlich das Komplementsystem. Dadurch werden Makrophagen und T-Helferzellen chemotaktisch zu der Stelle gelockt, wo das Antigen eindringt. Da diese Effekte zum Teil unspezifisch sind (z.B. die Anlockung der Makrophagen), kann eine in Gang gekommene Immunreaktion auch oft einen vorübergehenden Schutz gegen solche Mikroorganismen bieten, gegen die noch gar keine Immunität erworben wurde.
Immunreaktion gegen Viren
Viren können sich nur in den Wirtszellen vermehren. Da freie Antikörper nicht in Zellen eindringen, ist die Immunreaktion über Antikörper nur gegen solche Viren wirksam, die sich nicht in Zellen befinden. Werden Antikörper an die Virusoberfläche gebunden, so kommt durch Aktivierung des Komlementsystems eine Zerstörung der Viren in Gang. Häufig tragen Zellen, die von Viren befallen sind, in ihrer Zellmembran Komponenten, die von den betreffenden Viren stammen. T-Killer-Zellen erkennen diese Komponenten und töten die infizierten Zellen ab. Die Zerstörung körpereigener Zellen ist also bei einer Bekämpfung von Virusinfektionen ein normaler Vorgang. Die Aktivität der T-Killer-Zellen wird durch MHC-Proteine reguliert; diese haben also einen Einfluß darauf, wie gut das Immunsystem auf Virusinfektionen (z.B.: Grippeviren) reagieren kann.
Viele Viren besitzen eine große Variabilität in ihren Oberflächenstrukturen (sie liegen als verschiedene Stämme vor; dies gilt z.B.: für Grippe-, Schnupfen-, Maul-und Klauenseuche-Viren) oder sie verändern diese Strukturen relativ rasch und bilden neue Stämme (z.B.: das Grippevirus). Gegen derartige Viren ist die erworbene Immunität in der Regel viel weniger wirksam als gegen Bakterien, da bei jedem neuen Virusstamm eine zusätzliche Immunität erworben werden muß.
Immunreaktion gegen tierische Einzeller
Malaria, Amöbenruhr und Schlafkrankheiten werden von Einzellern hervorgerufen. Auch diese Organismen werden vom Körper durch Immunreaktionen bekämpft, die allerdings noch nicht völlig geklärt sind. Der Malariaerreger lebt lange Zeit intrazellulär (in Leberzellen bzw. Erythrozyten??) und verändert dabei die Zellmembran der Wirtszellen kaum. Daher erfolgt in dieser Zeit keine Immunreaktion. Trypanosomen (Erreger der Schlafkrankheit) entgehen der gegen sie gerichteten Immunreaktion teilweise dadurch, daß sie die als Antigene wirksamen Proteine ihrer Zellmembran rasch verändern. Eine zweite Infektion wird daher von den Gedächtniszellen vielfach nicht erkannt.
Allergische Reaktionen
Übermäßige oder ungeeignete Immunreaktionen führen zu Krankheits-erscheinungen, die man zusammenfassend als allergisch bezeichnet. Entsprechend der Komplexität der Immunreaktion kann die Störung an ganz unterschiedlichen Stellen eintreten. Daher gibt es sehr verschiedene allergische Reaktionen.
Anaphylaktische Reaktion
Bei einer ersten Reaktion eines Antigens bilden sich auch Antikörper der IgE-Klasse. Diese bilden sich an Mastzellen(??). Normalerweise verhindern T-Unterdrücker-Zellen, daß zu viele IgE-Antikörper gebildet und Mastzellen damit besetzt werden. Wenn die Unterdrücker-Zellen zu wenig wirksam sind, lagern sich zu viele IgE-Moleküle an Mastzellen an und es tritt eine Überempfindlichkeit (Allergie) gegnüber dem Antigen auf. Die ist vor allem bei sonst meist harmlosen Antigenen (Blütenstaub, Staub, Erdbeeren, Fischeiweiß, Arzneimittel) zu beobachten. Solche Allergien sind z.B.: Heufieber, Hautausschläge, allergisches Asthma. In manchen Fällen kann eine schwere Kreislaufstörung (anaphylaktischer Schock mit Blutdruckabfall, Schwäche, Pulsbeschleunigung) auftreten. Vor allem bei heftigen Allergien gegen Insektenstiche kann ein solcher Schock lebensgefährlich werden. Die anaphylaktische(??) Reaktion ist auf die Stimulierung übermäßig vieler Mastzellen zurückzuführen.
Autoimmun-Erkrankungen
Immunreaktionen können erst einige Tage bis Wochen nach der Geburt ablaufen. Erst dann ist die Entwicklung des Immunsystems abgeschlossen. Körperfremde Stoffe, die vorher in den Körper gelangen, lösen keine Immunreaktion aus; sie werden "geduldet" (Immuntoleranz). Die Unterscheidung körperfremder und körpereigener Stoffe "lernt" das Immunsystem bereits während der Embryonalzeit.
Mitunter werden Antikörper auch gegen körpereigene Gewebe gebildet; die führt zu Autoimmun-Krankheiten. Die Ursachen dafür sind nur teilweise bekannt. Manche Körperzellen besitzen normalerweise keine MHC-Proteine(??) in ihrer Zellmembran. Wenn solche Zellen infolge eines Fehlers im Zellgeschehen dann doch MHC-Proteine bilden, so reagieren T-Zellen darauf wie auf Antigene, obwohl es sich um körpereigene Zellbestandteile handelt. So wird eine Immunreaktion gegen diese Zellen ausgelöst. Eine bestimmte Form der Zuckerkrankheit (Diabetes mellitus), die vorwiegend bei Jugendlichen auftritt, ist eine solche Autoimmun-Erkrankung. In diesem Fall werden Inselzellen des Pankreas als körperfremd angesehen und abgebaut.
In anderen Fällen werden Membranproteine bestimmter Zellen als fremd angesehen.
Immunkomplex-Überreaktion
Die Immunkomplexe werden normalerweise rasch abgebaut. Tritt eine Verzögerung ein, so kommt es zu einer heftigen Entzündung, die bis zur Gewebsschädigung führen kann. In manchen Fällen werden dann Antikörper gegen die Immunkomplexe gebildet, sodaß Autoimmun-Reaktionen hinzukommen. Die Immunkomplexe können auch im Körper wandern und vielerorts Entzündungen auslösen. Dies ist z.B.: bei der Allergie gegen Penicillin der Fall; es kommt dann zu Nesselfieber (allgemeine Entzündung der Haut) und zu Gelenk- und Muskelschmerzen. Immunkomplexe vermögen sich an vielen Stellen des Körpers vorübergehend festzusetzen. Erfolgt dies in den Nieren-Glomeruli(??), so entsteht eine Nierenentzündung. In Gelenken verursachen Immunkomplexe arthritische Entzündungen, die oftmals im Körper "wandern". Auch andere rheumatische Erkrankungen gehen auf Immunkomplex-Überreaktion oder auf Autoimmun-Reaktion zurück.
Organverpflanzung; Abstossung von Fremdgewebe
Verpflanzt man ein Organ in einen fremden Körper, so bildet dieser Antikörper gegen das fremde Gewebe. Einige der Proteine des transplantierten Gewebes sind nämlich mit dem Immunsystem des eigenen Körpers nicht verträglich; sie wirken als Antigene. Vor allem sind dies MHC-Proteine, die dadurch ja ihren Namen erhalten haben. Wenn eine Reaktion der Gewebeverträglichkeits-Antigene mit Antikörpern eingetreten ist, werden die T-Killer-Lymphozyten tätig und zerstören das fremde Gewebe.
Die Verpflanzung von Organen von einem Körper in einen anderen ist dann erfolgreich, wenn beide genau die gleichen Gewebeverträglichkeits-Antigene besitzen, was bei der großen Variationsbreite fast nur bei eineiigen Zwillingen der Fall ist. In allen anderen Fällen tritt eine Immunreaktion ein. Je nach dem Grad der Übereinstimmung der MHC-Proteine von Spender und Empfänger fällt die Reaktion stärker oder schwächer aus. Man muß sie natürlich möglichst gering halten. Aus diesem Grunde müssen die MHC-Proteine von Spender und Empfänger wenigstens teilweise übereinstimmen. Darüber hinaus muß die Immunreaktion durch Stoffe unterdrückt werden, die Lymphozyten funktionsunfähig machen (immunsuppressive Stoffe). Dazu gehören zellteilungshemmende Stoffe, welche die Vermehrung der Lymphozyten verhindern, oder Corticosteroide (Hormone der Nebennierenrinde). Diese Substanzen führen allerdings zu einer Schwächung des ganzen Immunsystems. Spezifischer wirkt das aus einem Pilz gewonnene Cyclosporin, das die Aktivierung von T-Zellen hemmt, ohne die B-Zellen zu beeinflussen. Nicht jedes Organ hat bei Verpflanzung die gleiche Wirkung auf das Immunsystem des Empfängers; die Hornhaut des Auges und Gehörknöchelchen lösen bei einer Verpflanzung normalerweise keine Immunreaktion aus. Sie zeigen Immuntoleranz. Die Transplantation einer Niere ist immunologisch viel leichter zu beherrschen als eine Verpflanzung von Herz oder Leber.
Übersteht ein Mensch eine Infektionskrankheit, dann ist er in der Regel sein Leben lang vor einem neuen Ausbruch dieser Krankheit bewahrt. Gedächtniszellen gegen diese Krankheit bleiben also lange erhalten und geben dadurch Schutz vor einer neuen Ansteckung derselben Art; der Körper ist gegen die betreffende Krankheit "immun" geworden. Auf dieser Tatsache beruht die erstmals von dem englischen Arzt Jenner 1796 angewandte Schutzimpfung zur Immunisierung des Körpers.
Bei der aktiven Immunisierung regt man den Körper selbst auf eine für ihn ungefährliche Weise zur Bildung der Antikörper an. Dazu injiziert man abgetötete oder abgeschwächte Krankheitserreger. Diese rufen keine Krankheit hervor, veranlassen aber den Körper zu einer Immunreaktion. Tritt einige Zeit später eine natürliche Infektion durch den Erreger ein, erfolgt sofort eine heftige Reaktion, die den Erreger unschädlich macht. Da Gedächtniszellen sehr langlebig sind, wirkt eine aktive Immunisierung für längere Zeit vorbeugend.
Bei der passiven Immunisierung verlegt man die Bildung der Antikörper auf ein anderes Lebewesen und überträgt dessen Serum, in dem die Antikörper enthalten sind, in den Körper des Erkrankten. Die passive Immunisierung dient zur Heilung bereits ausgebrochener Infektionskrankheiten. Durch die von außen zugeführten Antikörper wird der Organismus in seinem Kampf gegen die Erreger unterstützt, bis genügend eigene Antikörper entstanden sind.
Zur Gewinnung von Diphtherieheilserum spritzt man, nach Emil von Behring (1894), Pferden mehrfach Diphtherieerreger ein, sodaß sich im Pferdeblut reichlich Antikörper dagegen bilden. Sie werden aus abgezapftem Blut gewonnen und längere Zeit steril aufbewahrt. Rechtzeitig einem Diphtheriekranken eingespritzt vermag das Heilserum in der Regel den vollen Ausbruch der Krankheit zu verhindern. Eine passive Schutzimpfung ist normalerweise nur wenige Wochen wirksam. Der menschliche Körper bildet nämlich gegen die Serumproteine des Pferdes seinerseits Antikörper. Diese fällen die fremden Antikörper bald aus. Wird ein zweites Mal Pferdeserum injiziert, fällt die Immunreaktion oft sehr heftig aus (anaphylaktischer Schock). Zur Vermeidung solcher Reaktionen verwendet man bei wiederholter Impfung, zum Beispiel gegen Wundstarrkrampf(??) ein Humanserum oder gereinigte Antikörper.
Auf der Bildung spezifischer Antikörper gegen artfremde Proteine beruht auch die Serumreaktion. Da jede Tierart arteigene Proteine besitzt, veranlaßt eingespritztes Serum von Fremdblut den Organismus zur Bildung von Antikörpern, die artfremde Proteine ausfällen. Bringt man im Reagenzglas Blutserum des Empfängerblutes mit dem zur Einspritzung verwendeten Serum zusammen., so kann mit die Ausfällung als Niederschlag unmittelbar beobachten. Die Antikörper sind nur gegen diejenigen Bestandteile das Blutes wirksam, welche ihre Bindung veranlaßt haben.
Die Serumreaktion dient in der Gerichtsmedizin zur Unterscheidung von Menschen- und Tierblut. Will man einen Blutfleck als Menschenblut nachweisen, löst man ihn in physiologischer Kochsalzlösung auf. Vorher hat man einem Kaninchen mehrmals menschliches Blutserum injiziert, so daß dessen Blut genügend Angikörper gegen menschliche Blutproteine enthält. Man entnimmt diesem Kaninchen Blut und gewinnt daraus das Serum. Diesem Serum setzt man den in physiologischer Kochsalzlösung aufgslösten Blutfleck zu. Entsteht ein Niederschlag, dann handelt es sich um Menschenblut, während das Ausbleiben des Niederschlags Menschenblut ausschließt.
Die Reaktion zwischen Antigen und Antikörper wird in der Biochemie wegen ihrer hohen Empfindlichkeit und Spezifität zur Identifikation von Proteinen verwendet. Es gibt mehrere Techniken. Die gebräuchlichste ist die Immundiffusions-Methode (Ouchterlony-Technik). Man gießt in eine Petrischale eine Agarschicht und stanzt drei Löcher aus. In eines der Löcher wird die Lösung eines bekannten Proteins (Antigen) gefüllt, in ein zweites Loch Serum eines Kaninchens, das man vorher gegen dieses Protein immunisiert hat. In das verbliebene Loch wird die zu testende Lösung gegeben. Die in den Lösungen enthaltenen Substanzen diffundieren in die Agarschicht. Treffen ein Antigen und ein zusammengehörender Antikörper zusammen, fällen sie sich aus. Deshalb entsteht zwischen Serum und der bekannten Proteinlösung allmählich eine sichtbare Bande des gefällten Antigen-Antikörper-Komplexes. Entsteht auch zwischen der Testlösung und dem Serum eine Bande, muss die Testlösung das gleiche Protein enthalten wie die Proteinlösung bekannter Zusammensetzung.
Werden Antikörper nach herkömmlichen Verfahren zur passiven Immunisierung oder für die Serumreaktion gewonnen, so erhält man stets ein Gemisch verschiedener Antikörper, weil verschiedene Bestandteile einer Bakterienmembran, einer Virushülle oder einer Proteinoberfläche als Antigene wirken. Deshalb werden unterschiedliche B-Zellen aktiviert. Sie bilden dann Plasmazellen und diese erzeugen jeweils die entsprechenden Antikörper. Es gibt jedoch ein Verfahren zur Herstellung größerer Mengen identischer Antikörper, das von jeweils einem einzigen Lymphozyten ausgeht. So erzeugte Antikörper nennt man monoklonal. (Ein Klon ist die aus einer Mutterzelle oder einem Mutterorganismus durch ungeschlechtliche Vermehrung hervorgegangene erbgleiche Nachkommenschaft.) Man läßt zunächst von einem Kaninchen durch Injektion des Antigens die Antikörper bilden. Dann isoliert man Antikörper bildende Zellen aus der Milz des Kaninchens und mischt diese mit Zellen aus einem Myelom (einer besonderen Tumorart), die unbegrenzte Teilungsfähigkeit haben. Bestimmte Chemikalien lösen in der Mischkultur Zellverschmelzungen aus. So entstehen Hybridzellen mit unbegrenztem Wachstum, die Antikörper erzeugen. Die Mischkultur bring man auf Nährmedien au denen nur die Hybridzellen wachsen. Anschließend kommenn die Hybridzellen einzeln in je einen Behälter mit Kulturflüssigkeit, wo sie sich vermehren. Jede Hybridzelle kann nur eine Sorte von Antikörper erzeugen. Einige Zellen jeder Kultur werden in flüssigen Stickstoff eingefroren; sie sind so fast unbegrenzt haltbar und können jederzeit wieder in Kultur übernommen werden.
Monoklonale Antikörper sind in der Medizin und Biochemie unentbehrliche Hilfsmittel. So kann man gegen zahlreiche Substanzen spezifische Antikörper erzeugen; Tumorzellen lassen sich infolge ihrer veränderten Zelloberfläche nachweisen; ebenso ist ein Nachweis von Viren schon in geringen Mengen möglich.
Mit monoklonalen Antikörpern können neue Impfverfahren eingeführt werden. Will man gegen ein bestimmtes Virus impfen, so läßt man von einer Maus Antikörper gegen dieses Virus erzeugen und gewinnt dann mit der geschilderten Methode monoklonale Antikörper. Diese läßt man bei anderen Mäusen durch Injektion als Antigene einwirken. So werden nun Antikörper gegen die Antikörper gebildet; diese haben dann als 'Negativ vom Negativ' teilweise ähnliche Moleküloberflächen wie das Virus. Nach Auswahl der richtigen Antikörper kann man diese beim Menschen injizieren. Sie verursachen nun ihrerseits als Antigene eine Bildung von weiteren Antikörpern. Diese weisen in bestimmten Bereichen der Moleküloberfläche eine Ahnlichkeit zu der viruserkennenden Oberfläche der ursprünglichen Virus-Antikörper auf und erkennen deshalb ebenfalls das Virus (aktive Immunisierung ohne Gefahr, eine Virusinfektion auszulösen).
Eine in der modernen Biologie häufig verwendetes Verfahren zur Mengenbestimmung von Stoffen, die als Antigene wirksam werden können, beruht ebenfalls auf der Anwendung monoklonaler Antikörper. Man bindet diese an einen Festkörper und läßt dann das Antigen einwirken. Außerdem werden gleichartige Antikörper-Moleküle mit einem Enzym verknüpft, dessen Reaktion leicht meßbar ist. Diese an das Enzym gekoppelte Antikörper läßt man mit dem am Festkörper gebundenen Antiköerper-Antigen-Komplex reagieren. Eine Reaktion erfolgt mir dort, wo das Antigen gebunden ist. Nun wird das Substrat des Enzyms zugesetzt und die Intensität der Enzymreaktion bestimmt; sie ist das Maß für die Menge gebundenen Antigens.
Bringt man Blut verschiedener Personen zusammen, so kann es sich entweder einfach vermischen oder es werden die Blutkörperchen zu kleinen Klumpen zusammengeballt. Als Ursache dieser Verballung (Agglutination) hat der Österreicher Landsteiner 1901 zwei verballungsfähige Stoffe A und B erkannt, die als Antigene wirken. Diese Blutgruppensubstanzen A und B sind Glykolipide und befinden sich in der Membran der Roten Blutkörperchen (Erythrozyten). Es gibt Menschen, deren Erythrozyten einen dieser Stoffe besitzen. Bei anderen kommen beide gemeinsam vor und bei wieder anderen fehlen beide. Die Zusammenballung selbst wird durch zwei verschiedenen Antikörper, Anti-A und Anti-B hervorgerufen; diese erkennen die Kohlenhydratketten der Glykolipide, die von der Zellmembran der Erythrozytenzellen nach außen ragen. Die Antikörper sind im Blutserum gelöst. Erythrozyten der Gruppe A werden nur von Anti-A, solche der Gruppe B nur von Anti-B verballt. Die Antikörper können daher im Serum nur dann vorhanden sein, wenn die entsprechenden Blutkörperchen fehlen, und umgekehrt. Nach der Verteilung dieser Stoffe hat man beim Menschen vier verschiedenen Blutgruppen aufgestellt. Angehörige der Blutgruppe A haben Erythrozyten mit dem Glykolipid A und im Blutserum den Antikörper Anti-B. Blutgruppe B besitzt Erythrozyten mit Glykolipid B und den Antikörpern Anti-A. Bei der Blutgruppe AB weisen die Erythrozyten beide Glykolipide, also A und B, auf; das Serum enthält keinen der Antikörper Anti-A oder Anti-B und kann deshalb auch übertragenes Blut nicht zum Verklumpen bringen. Die Erythrozyten der Blutgruppe 0 (null) haben weder Glykolipid A noch B. Das Serum dieser Blutgruppe enthält sowohl Anti-A als auch Anti-B und kann daher Blut jeder anderen Blutgruppe außer der eigenen zusammenballen.
Da Antikörper in nennenswerter Menge nur durch eine erste Immunreaktion entstehen, ist zu fragen, weshalb die Antikörper Anti-A und Anti-B auch ohne Berührung mit fremdem Blut vorhanden sind. Dies beruht darauf, daß die Antikörper auch gegen Oberflächenzuckerketten von Bakterien wirksam sind. Im ersten Lebensjahr erfolgt nach einer Infektion mit den entsprechenden Bakterien (vorwiegend solche der Darmflora) eine Bildung dieser Antikörper. Sie unterbleibt jedoch, wenn die Gegenwart der Kohlenhydratkette der Glykolipide A bzw. B in der Membran der Roten Blutzellen anzeigt, daß diese Kette als "körpereigen" anzusehen ist.
Außer den vier klassischen Blutgruppen, dem A/B/0-System, kennt man ein weiteres, für die Blutübertragung wichtiges Blutmerkmal /ein Antigen), den Rhesusfaktor. Er ist ebenfalls an die Oberfläche der Roten Blutkörperchen gebunden. Entdeckt wurde er 1940 im Blutserum von Kaninchen, das mit Blut von Rhesusaffen zur Antikörperbildung angeregt worden war. Menschen mit dem Rhesusfaktor-Antigen bezeichnet man als Rh-positiv (Rh), die übrigen als Rh-negativ (rh). Über 80% der Mitteleuropäer sind Rh-positiv. Ein Antikörper gegen das Antigen "Rhesusfaktor" bildet sich erst Monate nach einer Übertragung von Blut mit Rh-positiven Erythrozyten auf Rh-negative Menschen. Deshalb schadet einem Rh-negativen Menschen die erstmalige Übertragung von Blut mit Rh-positiven Erythrozyten nicht. Da die nach der Erstübertragung gebildeten Antikörper und Gedächtniszellen jedoch lange Zeit erhalten bleiben, können weitere Übertragungen von Blut mit Rh-positiven Erythrozyten zur Verballung und so zu schweren Schädigungen oder gar zum Tode führen. Schon im Mutterleib kann ein Kind davon betroffen sein.
Bei Rh-positiven Neugeborenen aus Beziehungen zwischen Rh-positiven Männern und Rh-negativen Frauen tritt zuweilen eine schwere Krankheit (Erythroblastose) auf. Bei der ersten Geburt können Erythrozyten des Rh-positiven Kindes in das mütterliche Blut mit Rh-negativen Erythrozyten übergehen und dort die Bildung von Antikörpern bewirken. Bei einer erneuten Schwangerschaft gelangen derartige Antikörper von der Mutter durch die Plazenta in das Blut des Kindes. Ist dieses ebenfalls Rh-positiv, so binden diese Antikörper an die Roten Blutkörperchen des Kindes und zerstören sie. Dadurch verringert sich der Sauerstofftransport und aus dem freigesetzten Hämoglobin entstehen gelb und grün gefärbte Abbaustoffe, welche die Leber schädigen und Gelbsucht hervorrufen. Weitere Rh-positive Kinder werden daher entweder tot geboren oder sind nur kurze Zeit lebensfähig. Verhindert man die Bildung von Antikörpern im Blut der Mutter, so tritt diese Auswirkung nicht ein. Man führt daher eine passive Immunisierung durch, indem man der Rh-negativen Mutter nach der Geburt des ersten Rh-positiven Kindes eine Serum mit Antikörpern gegen das Rhesusfaktor-Antigen injiziert. Diese Antikörper lagern sich an die eingedrungenen Blutzellen des Kindes an, die dann in der Mutter keine Bildung von Antikörpern mehr auslösen können. Die mit Antikörpern besetzten Blutzellen und die injizierten Antikörper werden abgebaut.
Die Blutgruppenzugehörigkeit und der Rhesusfaktor bleiben während des ganzen Lebens unverändert. Sie werden nach den Mendelschen Regeln vererbt. Blutgruppen sind bei allen menschlichen Rassen nachgewiesen worden.
Die Blutgruppenzugehörigkeit ist für die Blutübertragung von größter Bedeutung. Es wird grundsätzlich nur Blut von Angehörigen derselben Blutgruppe übertragen. Genauere Untersuchungen machen eine Aufteilung der Blutgruppe A in die Untergruppen A1 und A2, beim Rhesusfaktor in die Gruppen C, D und E notwendig. Ihre Häufigkeit in unserer Bevölkerung zeit Tabelle 186/1.
Bei den meisten Menschen werden die Blutgruppen-Substanzen auch in Sekreten ausgeschieden. Die Gerichtsmedizin kann deshalb in Blut-, Schweiß - und Speichelspuren die Blutgruppe nachweisen und dadurch bei Verbrechen Verdächtige ausschließen. Sie zieht dazu auch weitere Blutkörpercheneigenschaften heran, welche für die Blutübertragung wegen des Fehlens der Antikörper meist bedeutungslos sind. Die Blutgruppen spielen auch bei Abstammungsfragen etwa zur Klärung der Vaterschaft eines Kindes eine Rolle.
In vielen Fällen wird bei einer Blutübertragung kein Vollblut mehr verwendet. Statt dessen isoliert man aus dem Vollblut durch Zentrifugieren denjenigen Bestandteil, den der jeweilige Patient braucht und überträgt nur diesen (z.B. Rote Blutkörperchen oder Blutplättchen oder einen Gerinnungsfaktor).
Das menschliche Immunsystem kann mindestens zehn Millionen verschiedene Antikörper bilden. Da die Gesamtzahl der Gene beim Menschen sicher unter einer Million liegt, erhebt sich die Frage, wie die Vielfalt der Antikörper-Proteine entsteht. Ein vollständiger Antikörper besteht aus leichten und schweren Ketten und jede dieser Ketten aus einem konstanten und einem variablen Teil. Der variablen und der konstanten Region der Antikörper liegen jeweils eigene Gene zugrunde. Diese Gene liegen im Genom embryonaler Zellen von Säugern und der meisten Körperzellen weit voneinander entfernt, in den Antikörper bildenden Zellen hingegen eng benachbart.
Die Gene für die leichten und die schweren Ketten liegen auf verschiedenen Chromosomen; es handelt sich um getrennte Gengruppen. In jeder Zelle, die Antikörper bildet, ist aber stets nur je eines der Gene für eine leicht Kette und für einen schwere Kette aktiv, sodaß also nur eine Art von leichten Ketten und eine Art von schweren Ketten gebildet wird. Wie in allen Körperzellen ist wegen des doppelten Chromosomensatzes jedes Gen doppelt vorhanden; es wird jedoch immer nur ein mütterliches oder ein väterliches Allel eines Gens aktiviert. Wie dieser Allelen-Auschluß zustande kommt, ist unbekannt. Ein strenger Allelen-Ausschluß erfolgt sonst nur noch bei der Inaktivierung eines X-Chromosoms im weiblichen Geschlecht.
Jede Gengruppe für eine Immunglobulin-Kette besteht aus 100 bis 200 verschiedenen V-Genen für die variable Region und aus einem C-Gen (bei den schweren Ketten aus mehreren C-Genen) für die konstante Region. Während der Entwicklung der B-Zellen im Knochenmark werden die V-Gene in die Nachbarschaft eines C-Gens umgelagert. Dadurch wird bereits eine erhebliche Variabilität der Genkombination zwischen den beiden Genen erreicht. Erst nach der Gen-Umlagerung kann die jeweilige Genkombination aktiv werden.
Leichte und schwere Ketten treten zu einem Antikörpermolekül zusammen; somit erhält man mindestens 1000 x 10000 =10 hoch 7 verschiedenen Antigen bindende Oberflächen von Antikörpern. Jedoch sind damit die Variationsmöglichkeiten noch nicht erschöpft.
Im Bereich der V-Gene finden während der Ausbildung der Gedächtniszellen sehr häufig somatische Mutationen statt. Dabei kommt es zu der außerordentlich hohen Mutationsrate von 10³ Basenveränderungen je Zelle und Zellteilungsvorgang . Hierdurch wird die Variabilität der Antikörper um mehr als das Zehnfache weiter erhöht. Bei einer erneuten Immunreaktion erfolgt dann eine Auswahl solcher Zellen, deren Antikörper besonders gut reagieren (somatische Selektion), dies trägt wesentlich zu der verstärkten zweiten Immunreaktion bei.
Neu gebildete B-Zellen bilden zunächst stets IgM-Moleküle; später gehen sie zur Bildung von IgG-Molekülen über. Dazu muß ein anderes C-Gen aktiv werden, während das V-Gen-System unverändert bleibt. Die Spezifität des Antikörpers bleibt gleich, da sie nur durch den V-Bereich festgelegt ist. Daher haben Antikörper verschiedener Klassen, die nacheinander von der gleichen Zelle gebildet werden, immer die gleichen Bindungseigenschaften.
Antikörper und Gentechnik.
Die hohe Bindungsspezifität der Antikörper hofft man ausnutzen zu könne, indem man gentechnisch in Antikörper bestimmte Enzymeigenschaften einbaut, sodaß diese Enzyme dann ganz gezielt binden und ihre Wirkung entfalten. Ferner versucht man monoklonale Antikörper zu produzieren, deren konstante Regionen vom Menschen, die variablen Regionen aber von der Maus herstammen. Solchen "chimären Antikörper" würde das menschliche Immunsystem vermutlich viel weniger als "fremd" ansehen als rein tierische Antikörper. Daher sollten chimäre Antikörper für die passive Immunisierung vorteilhaft sein.
Die Gene der variablen Regionen von Maus-Antikörpern gegen menschliche Lymphozyten können gentechnsich in Zellkulturen in Gene von Antikörpern des Menschen eingebaut werden; die so gewonnenen "chimären Antikörper" können in Serumflüssigkeit vom Arzt eingesetzt werden, um menschliche Lymphozyten zu vernichten.
1986 entdeckte man, daß das 'human immunodeficiency virus', oder HIV für eine Krankheit des Immunsystems; das Erworbene Immunschwäche-Syndrom oder 'acquired immunodeficiency syndrome' kurz AIDS genannt, verantwortlich ist. AIDS ist das Endstadium eines langen infektiösen Prozesses, der klinisch durch eine symptomfreie Zeit gekennzeichnet ist, die Jahre andauern kann.
HIV ist ein gekapselter Retrovirus, der RNA als Genom enthält. Es gibt 2 Subtypen: HIV-1 und HIV-2. Der erste ist die Hauptursache von AIDS weltweit, HIV-2 wird hauptsächlich in westafrikanischen Ländern gefunden.
Haupt | Fügen Sie Referat | Kontakt | Impressum | Nutzungsbedingungen